|
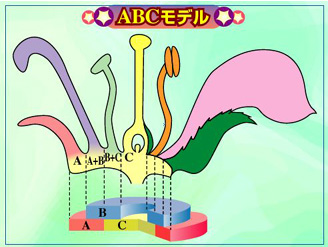
以上のような、既報の遺伝学的解析結果と新たな解剖結果を加味すると、変化アサガオの花の実際の変異は、表F271のように推定される。萼片の増加する変異(H、I、J)に関しては遺伝解析されたものはなく、解剖もされていないので、正確な花の構成はわからない。これらの変異のうち、(D)~(J)がホメオティック変異と思われる。(F)の変異はクラスC遺伝子の欠損としてABCモデルで説明できる。しかし、それ以外はABCモデルだけでは説明できない。
表F271.変化アサガオにおける花構成。( )内のアルファベットは図F231と表F241の記号と対応する。(A)は野生型。
(A)萼片-花弁-雄しべ-雌しべ
(B,C)萼片-花弁(増数)-雄しべ-雌しべ(増数)
(D,E)萼片-花弁-花弁-雌しべ
(F)萼片-花弁-花弁-<以上の繰り返し>
(G)萼片-花弁-花弁-シュート
(H)萼片(増数)-花弁(増数)-?
(I)萼片(増数)-萼片(増数)-花弁(増数)-?
(J)萼片(増数)-萼片(増数)-?
変化アサガオではABCモデルだけでは説明できない変異が多いと推測される。しかし、シロイヌナズナでも、ABCモデルどおりに花の形がデザインされた上で、別の変異が加わって、さらに多様な形態変異が生じる。例えば、雄しべ領域数が増えるシロイヌナズナsuperman変異体は、SUPERMAN遺伝子の影響でBクラス遺伝子が第4領域でも機能すると考えることで説明できる。すべての領域において花器官数が増加するclavata1変異体も同様に考えることができる。おそらく、アサガオでも、ABC遺伝子以外の未知の遺伝子の働きが加わって、多様な変化を生み出したのだろう。変化アサガオの研究から、花形態形成に関与する新たな遺伝子が見いだされる可能性がある。アサガオでは同じ領域の増加、領域内での器官の増加が多いので、SUPERMANやCLAVATA1のような遺伝子がみつかるかもしれない。
|